Benjamin
New member
- Joined
- Jun 12, 2007
- Messages
- 7
- Reaction score
- 3
- Points
- 0
- Location
- Jersey, Channel islands
- Country
- England
- Display Name
- Ben
Hi Guys
Sorry for the wait, this turned into a bit of a mammoth undertaking. I would like to thank everyone who responded to my plea for information on the forum, you were all a great help. For a PDF copy of the gudelines please follow this link to download http://www.sendspace.com/file/a3ma56
There is an appendix which I could not include here on the forum. The appendix has photos of a postmortem dissection of Typhlonectes natans
Husbandry Guidelines for
aquatic typhlonectid caecilians
(Typhlonectes sp.)

B. Tapley ©



Benjamin Tapley
Durrell Wildlife Conservation Trust
2009
Husbandry Guidelines for aquatic typhlonectid caecilians
(Typhlonectes spp.)
SECTION 1. BIOLOGY AND FIELD DATA
BIOLOGY
1.1 Taxonomy
ORDER: GYMNOPHIONA
FAMILY: Caeciliidae
GENUS: Typhlonectes
SPECIES: T. compressicauda
T. natans
COMMON NAMES
Typhlonectes sp. Rubber Eel, Caecilian worm, Aquatic caecilian.
T. natans: Rio Cauca Caecilian, Anguilla
T. compressicauda: Cayenne Caecilian, Cunha’s Caecilian
1.2 Morphology
LENGTH
T. natans, Adult size 250-725mm total length (Gymnophiona.org; Duellman & Trueb, 1994)
T. compressicauda 523mm Total length (Gymnophiona.org)
WEIGHT
T. natans may double their weight prior to giving birth, Kowalski reports an adult female T. natans weighing 250.0g in captivity.

Figure 1. Adult female T. natans B. Tapley ©
COLOURATION
T. natans are dark grey with a lighter ventral surface. (Figure 1) and T. compressicauda are dark grey to dark brown with a lighter ventral surface.
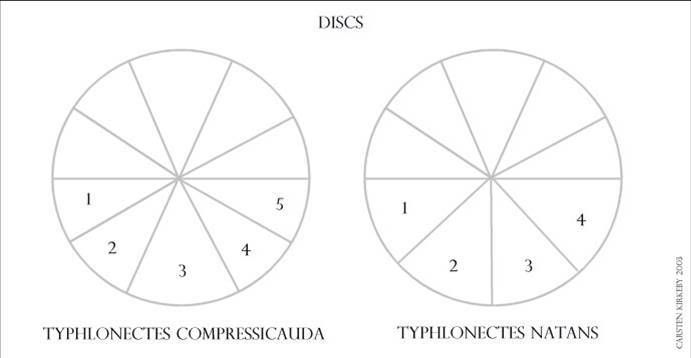
Figure 2. Cloacal discs showing the differences between T. natans and T. compressicauda.

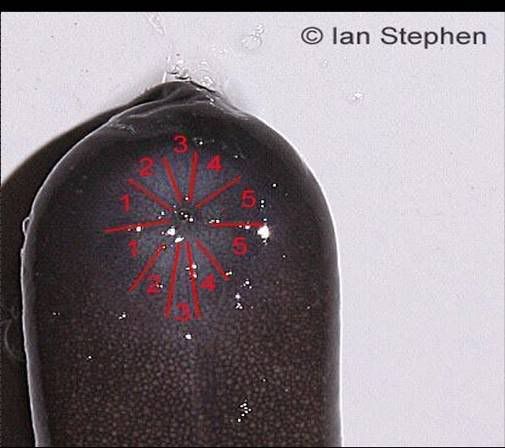
Figure 3. The cloacal disc in T. compressicauda.
DESCRIPTION
Externally it is difficult to tell T. natans and T. compressicauda apart. Typhlonectids are legless and have a wormlike body due to the presence of cutaneous folds (each fold is known as an annuli). The eyes are small and covered with skin and the mouth is recessed. There is a tentacle (for olfaction and tactile sensory purposes) located in the nasolabial groove. Small external nares are present. T. compressicauda has a laterally compressed dorsal fin situated along the caudal third of the body, this fin is absent or far less developed in T. natans. In typhlonectids the cloaca is situated on a cloacal disc at the terminus of the body (Wright and Whitaker, 2001). A tail is absent (Duellman & Trueb, 1994)
The only reliable way to tell T. natans from T. compressicauda is to examine the cloacal disc (Fig 3) The lower half of the cloacal disc (anterior part on the caecilians) is divided into 5 sections in T. compressicauda and into 4 sections in T. natans. The posterior half of the cloacal disc is divided into five sections in both species.
1.3 Physiology
RESPIRATORY RATE
Frequency of lung ventilation for T. natans 6.33 ±0.84 breaths hr -1 (Prabha et al., 2000)
GAS EXCHANGE
94% of gaseous exchange for T. compressicauda derived from pulmonary respiration (Sawaya, 1947)
1.4 Longevity
Maximum recorded age for T. natans, 13 years. Sp. still living
FIELD DATA
1.5 Zoogeography / Ecology
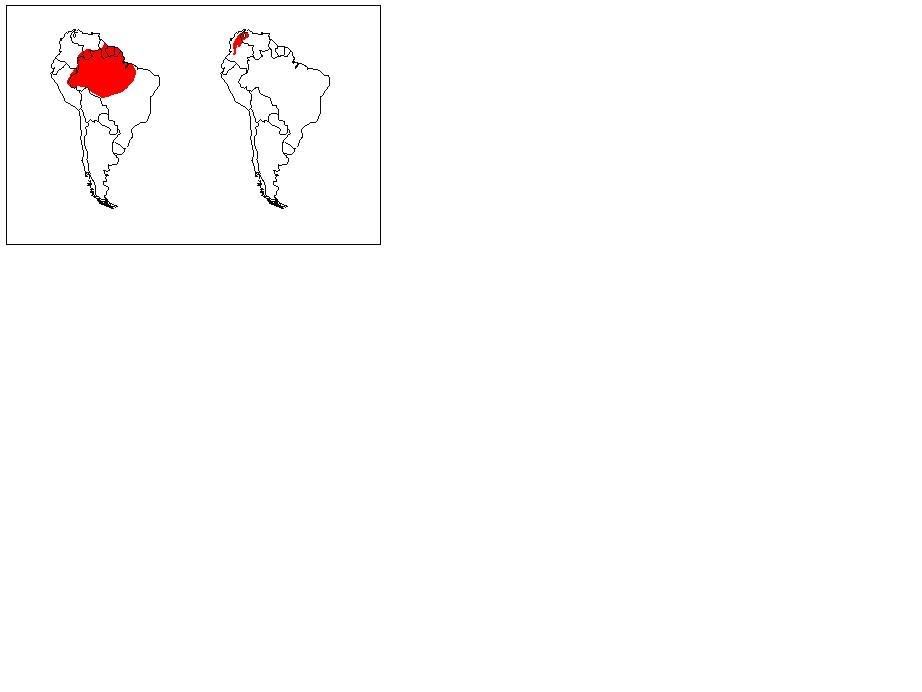
Figure 4. The distribution of T. compressicauda (left) and T. natans (Right)
DISTRIBUTION
T. natans - Cauca and Magdalena Drainages of Colombia and North eastern Venezuela (Fig. 4)
T. compressicauda - Amazon basin of Brazil, Colombia, French Guiana, Guyana, Peru, Surinam, Peru, and Venezuela (Fig. 4)
HABITAT
T. natans - Is an aquatic species, living in rivers, marshes and lakes, usually in open areas, and is only rarely found on land (IUCN et al, 2008). T. natans has been recorded in eutrophic waters in synanthropic and urbanized habitats in Venezuela (Gower & Wilkinson, 2005).
T. compressicauda - Is an aquatic species, inhabiting permanent rivers and marshes, mainly in the lowland forest zone (IUCN et al, 2008). Moodie (1978) reports that T. compressicauda inhabits water filled burrows.
CONSERVATION STATUS & POPULATION
T. natans - Least Concern in view of its wide distribution, tolerance of significant habitat degradation, presumed large population, and because it is unlikely to be declining fast enough to qualify for listing in a more threatened category (IUCN et al, 2008).
T. compressicauda - Least Concern in view of its wide distribution, presumed large population, and because it is unlikely to be declining fast enough to qualify for listing in a more threatened category (IUCN et al, 2008).
1.6 Diet and Feeding Behaviour
FOOD PREFERENCE
Hofer (2000) notes that typhlonectids can be particularly abundant around fishing villages where they have been observed feeding on the discarded entrails of fish.
T. compressicauda – In the wild juveniles of this species consume oligochaete worms, aquatic insects, terrestrial insects and anuran larva and eggs (Verdade et al, 2000). Gut contents of wild specimens also contained large amounts of detritus and plant matter which could have been ingested accidentally or indicate partial detritvorous habits (Verdade et al, 2000). T. compressicauda has also been observed feeding on dead fish caught in nets in French Guyana (Exbrayat & Delsol, 1985). In Manaus the stomach contents adult T. compressicauda included shrimp and arthropods, local fisherman reported that the species also feeds on small fish (Moodie, 1978)
Little is known about the wild diet of T. natans.
FEEDING
T. compressicauda - Carnivorous and generalistic. Juvenile wild specimens have been observed actively foraging in shallow water at night. Analysis of gut contents found that prey items associated with the bottom of the aquatic habitat as well as items associated with the surface (perhaps encountered when caecilians surface to breath) (Verdade et al, 2000)
1.7 Reproduction
DEVELOPMENTAL STAGES TO SEXUAL MATURITY
Viviparity is characteristic of typhlonectids. Development of the embryo is supported by yolk reserves (Wells, 2007). These yolk reserves are rapidly depleted. Young of T. compressicauda may increase their length six times by the time their yolk is fully absorbed (Stebbins & Cohen, 1995). Once the yolk is depleted the embryos emerge from their egg membranes and uncurl, embryos then align themselves lengthways in the oviduct (Pough et al, 2002). The larvae are sustained by a lipid rich secretion from the walls of the oviduct. Oviductal young have well developed dentition which is very different to the dentition of the adults (Wake, 1976) The larvae use these specialised foetal teeth to scrape the oviductal epithelium which stimulates the secretion of the nutrient substance (Wake, 1976). The larvae of typhlonectids develop very large, highly vascularised sac like gills (Fig. 5) which are probably used for gas exchange across the maternal uterine wall (Wells, 2007), oxygen affinity of foetal blood in T. compressicauda is higher than that of adult blood (Duellman & Trueb, 1994). It has been suggested that the gills may uptake nutrients. These gills are lost within several hours after birth (Parkinson, 2004). At birth juvenile T. compressicauda may measure 40% of the females total length (Stebbins & Cohen, 1995).
 Figure 5. T. natans with external gills Wendy © M Zac
Figure 5. T. natans with external gills Wendy © M Zac
AGE OF SEXUAL MATURITY
T. compressicauda attain sexual maturity at 16months (Exbrayat & Delsol, 1985).
The age at which T. natans reach sexually maturity in the wild is unknown. In captivity a female was recorded being sexually mature at approximately 4 years of age (Ilze Dunce pers. comm.)
SEASONALITY OF CYCLING
The reproductive biology of typhlonectids in the wild has not been studied in great detail. One study on T. compressicauda in French Guyana found that males have a yearly cycle of spermatogenesis which occurs from June – July, copulation occurs between February and May (Exbrayat & Delsol, 1985). Females have a biennial cycle. During the first year females are vitellogenic from October to February. This period is when ovulation occurs and gestation begins. Parturition occurs between July and October. Vitellogenesis occurs once again but very quickly. During the second period of reproduction Ooctytes are not mature and they degenerate becoming atretic follicles (Exbrayat & Delsol, 1985).
It is thought that the reproduction of T. compressicauda is related to seasonal variation in rainfall. During the wet season there is an increased supply of food so this is when spermatogenesis, copulation and gestation occur (Exbrayat & Delsol, 1985).
T. natans is a seasonal breeder, breeding occurs during the wet season (Herman, 1994).
GESTATION PERIOD
In the wild gestation in T. compressicauda lasts six months (Exbrayat & Delsol, 1985).
In T. natans gestation period is unknown
NUMBER OF OFFSPRING
In the wild T. compressicauda give birth to on average 4 young. In captivity an average of 3 young are born (Exbrayat & Delsol, 1985).
The number of offspring produced by T. natans in the wild is unknown.
BIRTH DETAILS AND SEASONS
Young of T. compressicauda are born at the beginning of the dry season when water levels are still high (Exbrayat & Delsol, 1985).
Birth details and seasons of T. natans in the wild are unknown.
1.8 Behaviour
ACTIVITY
Activity patterns of typhlonectids in the wild are largely unknown. However T. compressicauda has been observed foraging in shallow water at night (Verdade et al, 2000)
LOCOMOTION
Lateral undulation.
PREDATION
Not known
VOCALISATION
In captivity a squeaking sound has been reported in Typhlonectids when they surface to breath (Stebbins & Cohen, 1995).
SOCIAL BEHAVIOUR
In captivity typhlonectids are gregarious and share refugia. Refugia sites appear to be marked with chemical signals, and individuals are attracted to chemicals emanated by conspecifics (Wells, 2007). A captive study on T. natans showed breeding males in captivity may be aggressive towards one another but not necessarily territorial (Wells, 2007). Tests show that non reproductive females preferred chemical cues from other females over male chemical cues. Reproductive males preferred cues from receptive females to unreceptive females and did not discriminate between related and unrelated females (Warbeck & Parzefall, 2001)
SEXUAL BEHAVIOUR
There are not many records of mating in Caecilians. One observation of T. natans showed that males entwine themselves around females, the lower bodies entwined vent to vent. Fertilisation is internal; the males then inserted the phallodeum, the intermittent organ of the caecilian (Fig. 6) into the cloaca of the female. Mating initially is frenzied but soon subsides. The animals can remain entwined for several hours (Parkinson, 2001). Murphy (1978) noted that copulation continued for three hours in T. compressicauda after it was first noted.
 Figure 6. A preserved male Typhlonectes natans illustrating protrusible phallodeum B. Tapley ©
Figure 6. A preserved male Typhlonectes natans illustrating protrusible phallodeum B. Tapley ©
Sorry for the wait, this turned into a bit of a mammoth undertaking. I would like to thank everyone who responded to my plea for information on the forum, you were all a great help. For a PDF copy of the gudelines please follow this link to download http://www.sendspace.com/file/a3ma56
There is an appendix which I could not include here on the forum. The appendix has photos of a postmortem dissection of Typhlonectes natans
Husbandry Guidelines for
aquatic typhlonectid caecilians
(Typhlonectes sp.)
B. Tapley ©
Benjamin Tapley
Durrell Wildlife Conservation Trust
2009
Husbandry Guidelines for aquatic typhlonectid caecilians
(Typhlonectes spp.)
SECTION 1. BIOLOGY AND FIELD DATA
BIOLOGY
1.1 Taxonomy
ORDER: GYMNOPHIONA
FAMILY: Caeciliidae
GENUS: Typhlonectes
SPECIES: T. compressicauda
T. natans
COMMON NAMES
Typhlonectes sp. Rubber Eel, Caecilian worm, Aquatic caecilian.
T. natans: Rio Cauca Caecilian, Anguilla
T. compressicauda: Cayenne Caecilian, Cunha’s Caecilian
1.2 Morphology
LENGTH
T. natans, Adult size 250-725mm total length (Gymnophiona.org; Duellman & Trueb, 1994)
T. compressicauda 523mm Total length (Gymnophiona.org)
WEIGHT
T. natans may double their weight prior to giving birth, Kowalski reports an adult female T. natans weighing 250.0g in captivity.
Figure 1. Adult female T. natans B. Tapley ©
COLOURATION
T. natans are dark grey with a lighter ventral surface. (Figure 1) and T. compressicauda are dark grey to dark brown with a lighter ventral surface.
Figure 2. Cloacal discs showing the differences between T. natans and T. compressicauda.
Figure 3. The cloacal disc in T. compressicauda.
DESCRIPTION
Externally it is difficult to tell T. natans and T. compressicauda apart. Typhlonectids are legless and have a wormlike body due to the presence of cutaneous folds (each fold is known as an annuli). The eyes are small and covered with skin and the mouth is recessed. There is a tentacle (for olfaction and tactile sensory purposes) located in the nasolabial groove. Small external nares are present. T. compressicauda has a laterally compressed dorsal fin situated along the caudal third of the body, this fin is absent or far less developed in T. natans. In typhlonectids the cloaca is situated on a cloacal disc at the terminus of the body (Wright and Whitaker, 2001). A tail is absent (Duellman & Trueb, 1994)
The only reliable way to tell T. natans from T. compressicauda is to examine the cloacal disc (Fig 3) The lower half of the cloacal disc (anterior part on the caecilians) is divided into 5 sections in T. compressicauda and into 4 sections in T. natans. The posterior half of the cloacal disc is divided into five sections in both species.
1.3 Physiology
RESPIRATORY RATE
Frequency of lung ventilation for T. natans 6.33 ±0.84 breaths hr -1 (Prabha et al., 2000)
GAS EXCHANGE
94% of gaseous exchange for T. compressicauda derived from pulmonary respiration (Sawaya, 1947)
1.4 Longevity
Maximum recorded age for T. natans, 13 years. Sp. still living
FIELD DATA
1.5 Zoogeography / Ecology
Figure 4. The distribution of T. compressicauda (left) and T. natans (Right)
DISTRIBUTION
T. natans - Cauca and Magdalena Drainages of Colombia and North eastern Venezuela (Fig. 4)
T. compressicauda - Amazon basin of Brazil, Colombia, French Guiana, Guyana, Peru, Surinam, Peru, and Venezuela (Fig. 4)
HABITAT
T. natans - Is an aquatic species, living in rivers, marshes and lakes, usually in open areas, and is only rarely found on land (IUCN et al, 2008). T. natans has been recorded in eutrophic waters in synanthropic and urbanized habitats in Venezuela (Gower & Wilkinson, 2005).
T. compressicauda - Is an aquatic species, inhabiting permanent rivers and marshes, mainly in the lowland forest zone (IUCN et al, 2008). Moodie (1978) reports that T. compressicauda inhabits water filled burrows.
CONSERVATION STATUS & POPULATION
T. natans - Least Concern in view of its wide distribution, tolerance of significant habitat degradation, presumed large population, and because it is unlikely to be declining fast enough to qualify for listing in a more threatened category (IUCN et al, 2008).
T. compressicauda - Least Concern in view of its wide distribution, presumed large population, and because it is unlikely to be declining fast enough to qualify for listing in a more threatened category (IUCN et al, 2008).
1.6 Diet and Feeding Behaviour
FOOD PREFERENCE
Hofer (2000) notes that typhlonectids can be particularly abundant around fishing villages where they have been observed feeding on the discarded entrails of fish.
T. compressicauda – In the wild juveniles of this species consume oligochaete worms, aquatic insects, terrestrial insects and anuran larva and eggs (Verdade et al, 2000). Gut contents of wild specimens also contained large amounts of detritus and plant matter which could have been ingested accidentally or indicate partial detritvorous habits (Verdade et al, 2000). T. compressicauda has also been observed feeding on dead fish caught in nets in French Guyana (Exbrayat & Delsol, 1985). In Manaus the stomach contents adult T. compressicauda included shrimp and arthropods, local fisherman reported that the species also feeds on small fish (Moodie, 1978)
Little is known about the wild diet of T. natans.
FEEDING
T. compressicauda - Carnivorous and generalistic. Juvenile wild specimens have been observed actively foraging in shallow water at night. Analysis of gut contents found that prey items associated with the bottom of the aquatic habitat as well as items associated with the surface (perhaps encountered when caecilians surface to breath) (Verdade et al, 2000)
1.7 Reproduction
DEVELOPMENTAL STAGES TO SEXUAL MATURITY
Viviparity is characteristic of typhlonectids. Development of the embryo is supported by yolk reserves (Wells, 2007). These yolk reserves are rapidly depleted. Young of T. compressicauda may increase their length six times by the time their yolk is fully absorbed (Stebbins & Cohen, 1995). Once the yolk is depleted the embryos emerge from their egg membranes and uncurl, embryos then align themselves lengthways in the oviduct (Pough et al, 2002). The larvae are sustained by a lipid rich secretion from the walls of the oviduct. Oviductal young have well developed dentition which is very different to the dentition of the adults (Wake, 1976) The larvae use these specialised foetal teeth to scrape the oviductal epithelium which stimulates the secretion of the nutrient substance (Wake, 1976). The larvae of typhlonectids develop very large, highly vascularised sac like gills (Fig. 5) which are probably used for gas exchange across the maternal uterine wall (Wells, 2007), oxygen affinity of foetal blood in T. compressicauda is higher than that of adult blood (Duellman & Trueb, 1994). It has been suggested that the gills may uptake nutrients. These gills are lost within several hours after birth (Parkinson, 2004). At birth juvenile T. compressicauda may measure 40% of the females total length (Stebbins & Cohen, 1995).
AGE OF SEXUAL MATURITY
T. compressicauda attain sexual maturity at 16months (Exbrayat & Delsol, 1985).
The age at which T. natans reach sexually maturity in the wild is unknown. In captivity a female was recorded being sexually mature at approximately 4 years of age (Ilze Dunce pers. comm.)
SEASONALITY OF CYCLING
The reproductive biology of typhlonectids in the wild has not been studied in great detail. One study on T. compressicauda in French Guyana found that males have a yearly cycle of spermatogenesis which occurs from June – July, copulation occurs between February and May (Exbrayat & Delsol, 1985). Females have a biennial cycle. During the first year females are vitellogenic from October to February. This period is when ovulation occurs and gestation begins. Parturition occurs between July and October. Vitellogenesis occurs once again but very quickly. During the second period of reproduction Ooctytes are not mature and they degenerate becoming atretic follicles (Exbrayat & Delsol, 1985).
It is thought that the reproduction of T. compressicauda is related to seasonal variation in rainfall. During the wet season there is an increased supply of food so this is when spermatogenesis, copulation and gestation occur (Exbrayat & Delsol, 1985).
T. natans is a seasonal breeder, breeding occurs during the wet season (Herman, 1994).
GESTATION PERIOD
In the wild gestation in T. compressicauda lasts six months (Exbrayat & Delsol, 1985).
In T. natans gestation period is unknown
NUMBER OF OFFSPRING
In the wild T. compressicauda give birth to on average 4 young. In captivity an average of 3 young are born (Exbrayat & Delsol, 1985).
The number of offspring produced by T. natans in the wild is unknown.
BIRTH DETAILS AND SEASONS
Young of T. compressicauda are born at the beginning of the dry season when water levels are still high (Exbrayat & Delsol, 1985).
Birth details and seasons of T. natans in the wild are unknown.
1.8 Behaviour
ACTIVITY
Activity patterns of typhlonectids in the wild are largely unknown. However T. compressicauda has been observed foraging in shallow water at night (Verdade et al, 2000)
LOCOMOTION
Lateral undulation.
PREDATION
Not known
VOCALISATION
In captivity a squeaking sound has been reported in Typhlonectids when they surface to breath (Stebbins & Cohen, 1995).
SOCIAL BEHAVIOUR
In captivity typhlonectids are gregarious and share refugia. Refugia sites appear to be marked with chemical signals, and individuals are attracted to chemicals emanated by conspecifics (Wells, 2007). A captive study on T. natans showed breeding males in captivity may be aggressive towards one another but not necessarily territorial (Wells, 2007). Tests show that non reproductive females preferred chemical cues from other females over male chemical cues. Reproductive males preferred cues from receptive females to unreceptive females and did not discriminate between related and unrelated females (Warbeck & Parzefall, 2001)
SEXUAL BEHAVIOUR
There are not many records of mating in Caecilians. One observation of T. natans showed that males entwine themselves around females, the lower bodies entwined vent to vent. Fertilisation is internal; the males then inserted the phallodeum, the intermittent organ of the caecilian (Fig. 6) into the cloaca of the female. Mating initially is frenzied but soon subsides. The animals can remain entwined for several hours (Parkinson, 2001). Murphy (1978) noted that copulation continued for three hours in T. compressicauda after it was first noted.
Last edited by a moderator:


")
